 Azərbaycanca (AZ)
Azərbaycanca (AZ) Deutsch (DE)
Deutsch (DE) English (US)
English (US) Қазақша (KZ)
Қазақша (KZ) Lietuva (LT)
Lietuva (LT) සිංහල (LK)
සිංහල (LK) Türkçe (TR)
Türkçe (TR) O'zbekcha (UZ)
O'zbekcha (UZ) 中國人 (CN)
中國人 (CN)The retina (from Latin rete 'net'; pl. retinae or retinas) is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.
| Retina | |
|---|---|
 Right human eye cross-sectional view; eyes vary significantly among animals. | |
| Details | |
| Pronunciation | UK: /ˈrɛtɪnə/, US: /ˈrɛtənə/, pl. retinae /-ni/ |
| Part of | Eye |
| System | Visual system |
| Artery | Central retinal artery |
| Identifiers | |
| Latin | rēte, tunica interna bulbi |
| MeSH | D012160 |
| TA98 | A15.2.04.002 |
| TA2 | 6776 |
| FMA | 58301 |
| Anatomical terminology [edit on Wikidata] | |
The neural retina consists of several layers of neurons interconnected by synapses and is supported by an outer layer of pigmented epithelial cells. The primary light-sensing cells in the retina are the photoreceptor cells, which are of two types: rods and cones. Rods function mainly in dim light and provide monochromatic vision. Cones function in well-lit conditions and are responsible for the perception of colour through the use of a range of opsins, as well as high-acuity vision used for tasks such as reading. A third type of light-sensing cell, the photosensitive ganglion cell, is important for entrainment of circadian rhythms and reflexive responses such as the pupillary light reflex.
Light striking the retina initiates a cascade of chemical and electrical events that ultimately trigger nerve impulses that are sent to various visual centres of the brain through the fibres of the optic nerve. Neural signals from the rods and cones undergo processing by other neurons, whose output takes the form of action potentials in retinal ganglion cells whose axons form the optic nerve.
In vertebrate embryonic development, the retina and the optic nerve originate as outgrowths of the developing brain, specifically the embryonic diencephalon; thus, the retina is considered part of the central nervous system (CNS) and is actually brain tissue. It is the only part of the CNS that can be visualized noninvasively. Like most of the brain, the retina is isolated from the vascular system by the blood–brain barrier. The retina is the part of the body with the greatest continuous energy demand.
Structure
Inverted versus non-inverted retina
The vertebrate retina is inverted in the sense that the light-sensing cells are in the back of the retina, so that light has to pass through layers of neurons and capillaries before it reaches the photosensitive sections of the rods and cones. The ganglion cells, whose axons form the optic nerve, are at the front of the retina; therefore, the optic nerve must cross through the retina en route to the brain. No photoreceptors are in this region, giving rise to the blind spot. In contrast, in the cephalopod retina, the photoreceptors are in front, with processing neurons and capillaries behind them. Because of this, cephalopods do not have a blind spot.
Although the overlying neural tissue is partly transparent, and the accompanying glial cells have been shown to act as fibre-optic channels to transport photons directly to the photoreceptors, light scattering does occur. Some vertebrates, including humans, have an area of the central retina adapted for high-acuity vision. This area, termed the fovea centralis, is avascular (does not have blood vessels), and has minimal neural tissue in front of the photoreceptors, thereby minimizing light scattering.
The cephalopods have a non-inverted retina, which is comparable in resolving power to the eyes of many vertebrates. Squid eyes do not have an analog of the vertebrate retinal pigment epithelium (RPE). Although their photoreceptors contain a protein, retinochrome, that recycles retinal and replicates one of the functions of the vertebrate RPE, cephalopod photoreceptors are likely not maintained as well as in vertebrates, and that as a result, the useful lifetime of photoreceptors in invertebrates is much shorter than in vertebrates. Having easily replaced stalk eyes (some lobsters) or retinae (some spiders, such as Deinopis) rarely occurs.
The cephalopod retina does not originate as an outgrowth of the brain, as the vertebrate one does. This difference suggests that vertebrate and cephalopod eyes are not homologous, but have evolved separately. From an evolutionary perspective, a more complex structure such as the inverted retina can generally come about as a consequence of two alternate processes - an advantageous "good" compromise between competing functional limitations, or as a historical maladaptive relic of the convoluted path of organ evolution and transformation. Vision is an important adaptation in higher vertebrates.
A third view of the "inverted" vertebrate eye is that it combines two benefits - the maintenance of the photoreceptors mentioned above, and the reduction in light intensity necessary to avoid blinding the photoreceptors, which are based on the extremely sensitive eyes of the ancestors of modern hagfish (fish that live in very deep, dark water).
A recent study on the evolutionary purpose for the inverted retina structure from the APS (American Physical Society) says that "The directional of glial cells helps increase the clarity of human vision. But we also noticed something rather curious: the colours that best passed through the glial cells were green to red, which the eye needs most for daytime vision. The eye usually receives too much blue—and thus has fewer blue-sensitive cones.
Further computer simulations showed that green and red are concentrated five to ten times more by the glial cells, and into their respective cones, than blue light. Instead, excess blue light gets scattered to the surrounding rods. This optimization is such that color vision during the day is enhanced, while night-time vision suffers very little".
Retinal layers




The vertebrate retina has 10 distinct layers. From closest to farthest from the vitreous body:
- Inner limiting membrane – basement membrane elaborated by Müller cells
- Nerve fiber layer – axons of the ganglion cell bodies (a thin layer of Müller cell footplates exists between this layer and the inner limiting membrane)
- Ganglion cell layer – contains nuclei of ganglion cells, the axons of which become the optic nerve fibres, and some displaced amacrine cells
- Inner plexiform layer – contains the synapse between the bipolar cell axons and the dendrites of the ganglion and amacrine cells
- Inner nuclear layer – contains the nuclei and surrounding cell bodies (perikarya) of the amacrine cells, bipolar cells, and horizontal cells
- Outer plexiform layer – projections of rods and cones ending in the rod spherule and cone pedicle, respectively, these make synapses with dendrites of bipolar cells and horizontal cells. In the macular region, this is known as the fiber layer of Henle.
- Outer nuclear layer – cell bodies of rods and cones
- External limiting membrane – layer that separates the inner segment portions of the photoreceptors from their cell nuclei
- Inner segment / outer segment layer – inner segments and outer segments of rods and cones, the outer segments contain a highly specialized light-sensing apparatus.
- Retinal pigment epithelium – single layer of cuboidal epithelial cells (with extrusions not shown in diagram). This layer is closest to the choroid, and provides nourishment and supportive functions to the neural retina, The black pigment melanin in the pigment layer prevents light reflection throughout the globe of the eyeball; this is extremely important for clear vision.
These layers can be grouped into four main processing stages—photoreception; transmission to bipolar cells; transmission to ganglion cells, which also contain photoreceptors, the photosensitive ganglion cells; and transmission along the optic nerve. At each synaptic stage, horizontal and amacrine cells also are laterally connected.
The optic nerve is a central tract of many axons of ganglion cells connecting primarily to the lateral geniculate body, a visual relay station in the diencephalon (the rear of the forebrain). It also projects to the superior colliculus, the suprachiasmatic nucleus, and the nucleus of the optic tract. It passes through the other layers, creating the optic disc in primates.
Additional structures, not directly associated with vision, are found as outgrowths of the retina in some vertebrate groups. In birds, the pecten is a vascular structure of complex shape that projects from the retina into the vitreous humour; it supplies oxygen and nutrients to the eye, and may also aid in vision. Reptiles have a similar, but much simpler, structure.
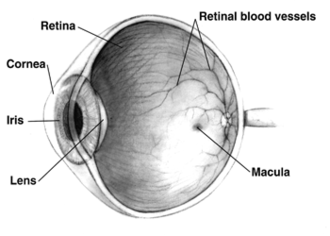
In adult humans, the entire retina is about 72% of a sphere about 22 mm in diameter. The entire retina contains about 7 million cones and 75 to 150 million rods. The optic disc, a part of the retina sometimes called "the blind spot" because it lacks photoreceptors, is located at the optic papilla, where the optic-nerve fibres leave the eye. It appears as an oval white area of 3 mm2. Temporal (in the direction of the temples) to this disc is the macula, at whose centre is the fovea, a pit that is responsible for sharp central vision, but is actually less sensitive to light because of its lack of rods. Human and non-human primates possess one fovea, as opposed to certain bird species, such as hawks, that are bifoviate, and dogs and cats, that possess no fovea, but a central band known as the visual streak.[citation needed] Around the fovea extends the central retina for about 6 mm and then the peripheral retina. The farthest edge of the retina is defined by the ora serrata. The distance from one ora to the other (or macula), the most sensitive area along the horizontal meridian, is about 32 mm.[clarification needed]
In section, the retina is no more than 0.5 mm thick. It has three layers of nerve cells and two of synapses, including the unique ribbon synapse. The optic nerve carries the ganglion-cell axons to the brain, and the blood vessels that supply the retina. The ganglion cells lie innermost in the eye while the photoreceptive cells lie beyond. Because of this counter-intuitive arrangement, light must first pass through and around the ganglion cells and through the thickness of the retina, (including its capillary vessels, not shown) before reaching the rods and cones. Light is absorbed by the retinal pigment epithelium or the choroid (both of which are opaque).
The white blood cells in the capillaries in front of the photoreceptors can be perceived as tiny bright moving dots when looking into blue light. This is known as the blue field entoptic phenomenon (or Scheerer's phenomenon).
Between the ganglion-cell layer and the rods and cones are two layers of neuropils, where synaptic contacts are made. The neuropil layers are the outer plexiform layer and the inner plexiform layer. In the outer neuropil layer, the rods and cones connect to the vertically running bipolar cells, and the horizontally oriented horizontal cells connect to ganglion cells.
The central retina predominantly contains cones, while the peripheral retina predominantly contains rods. In total, the retina has about seven million cones and a hundred million rods. At the centre of the macula is the foveal pit where the cones are narrow and long, and arranged in a hexagonal mosaic, the most dense, in contradistinction to the much fatter cones located more peripherally in the retina. At the foveal pit, the other retinal layers are displaced, before building up along the foveal slope until the rim of the fovea, or parafovea, is reached, which is the thickest portion of the retina. The macula has a yellow pigmentation, from screening pigments, and is known as the macula lutea. The area directly surrounding the fovea has the highest density of rods converging on single bipolar cells. Since its cones have a much lesser convergence of signals, the fovea allows for the sharpest vision the eye can attain.
Though the rod and cones are a mosaic of sorts, transmission from receptors, to bipolars, to ganglion cells is not direct. Since about 150 million receptors and only 1 million optic nerve fibres exist, convergence and thus mixing of signals must occur. Moreover, the horizontal action of the horizontal and amacrine cells can allow one area of the retina to control another (e.g. one stimulus inhibiting another). This inhibition is key to lessening the sum of messages sent to the higher regions of the brain. In some lower vertebrates (e.g. the pigeon), control of messages is "centrifugal" – that is, one layer can control another, or higher regions of the brain can drive the retinal nerve cells, but in primates, this does not occur.
Layers imagable with optical coherence tomography
Using optical coherence tomography (OCT), at least 13 layers can be identified in the retina. The layers and anatomical correlation are:


From innermost to outermost, the layers identifiable by OCT are as follows:
| # | OCT layer / Conventional label | Anatomical correlate | Reflectivity on OCT | Specific anatomical boundaries? | Ref. |
|---|---|---|---|---|---|
| 1 | Posterior cortical vitreous | Posterior cortical vitreous | Hyper-reflective | Yes | |
| 2 | Preretinal space | In eyes where the vitreous has fully or partially detached from the retina, this is the space created between the posterior cortical vitreous face and the internal limiting membrane of the retina. | Hypo-reflective | ||
| 3 | Internal limiting membrane (ILM) | Formed by Müller cell endfeet (unclear if it can be observed on OCT) | Hyper-reflective | No | |
| Nerve fiber layer (NFL) | Ganglion cell axons travelling towards the optic nerve | ||||
| 4 | Ganglion cell layer (GCL) | Ganglion cell bodies (and some displaced amacrine cells) | Hypo-reflective | ||
| 5 | Inner plexiform layer (IPL) | Synapses between bipolar, amacrine and ganglion cells | Hyper-reflective | ||
| 6 | Inner nuclear layer (INL) | a) Horizontal, bipolar and amacrine cell bodies b) Müller cell nuclei | Hypo-reflective | ||
| 7 | Outer plexiform layer (OPL) | Synapses between photoreceptor, bipolar and horizontal cells | Hyper-reflective | ||
| 8 | (Inner half) Henle's nerve fiber layer (HL) | Photoreceptor axons (obliquely orientated fibres; not present in mid-peripheral or peripheral retina) | Hypo-reflective | No | |
| (Outer half) Outer nuclear layer (ONL) | The photoreceptor cell bodies | ||||
| 9 | External limiting membrane (ELM) | Made of zonulae adherens between Müller cells and photoreceptor inner segments | Hyper-reflective | ||
| 10 | Myoid zone (MZ) | The innermost portion of the photoreceptor inner segment (IS) containing:
| Hypo-reflective | No | |
| 11 | Ellipsoid zone (EZ) | The outermost portion of the photoreceptor inner segment (IS) packed with mitochondria | Very Hyper-reflective | No | |
| IS/OS junction or Photoreceptor integrity line (PIL) | The photoreceptor connecting cilia which bridge the inner and outer segments of the photoreceptor cells. | ||||
| 12 | Photoreceptor outer segments (OS) | The photoreceptor outer segments (OS) which contain disks filled with opsin, the molecule that absorbs photons. | Hypo-reflective | ||
| 13 | Interdigitation zone (IZ) | Apices of the RPE cells which encase part of the cone OSs. Poorly distinguishable from RPE. Previously: "cone outer segment tips line" (COST) | Hyper-reflective | No | |
| 14 | RPE/Bruch's complex | RPE phagosome zone | Very Hyper-reflective | No | |
| RPE melanosome zone | Hypo-reflective | ||||
| RPE mitochondria zone + Junction between the RPE & Bruch's membrane | Very Hyper-reflective | ||||
| 15 | Choriocapillaris | Thin layer of moderate reflectivity in inner choroid | No | ||
| 16 | Sattler's layer | Thick layer of round or ovalshaped hyperreflective profiles, with hyporeflective cores in mid-choroid | |||
| 17 | Haller's layer | Thick layer of oval-shaped hyperreflective profiles, with hyporeflective cores in outer choroid | |||
| 18 | Choroidal-scleral juncture | Zone at the outer choroid with a marked change in texture, in which large circular or ovoid profiles abut a homogenous region of variable reflectivity | |||
Development
Retinal development begins with the establishment of the eye fields mediated by the SHH and SIX3 proteins, with subsequent development of the optic vesicles regulated by the PAX6 and LHX2 proteins. The role of Pax6 in eye development was elegantly demonstrated by Walter Gehring and colleagues, who showed that ectopic expression of Pax6 can lead to eye formation on Drosophila antennae, wings, and legs. The optic vesicle gives rise to three structures: the neural retina, the retinal pigmented epithelium, and the optic stalk. The neural retina contains the retinal progenitor cells (RPCs) that give rise to the seven cell types of the retina. Differentiation begins with the retinal ganglion cells and concludes with production of the Muller glia. Although each cell type differentiates from the RPCs in a sequential order, there is considerable overlap in the timing of when individual cell types differentiate. The cues that determine a RPC daughter cell fate are coded by multiple transcription factor families including the bHLH and homeodomain factors.
In addition to guiding cell fate determination, cues exist in the retina to determine the dorsal-ventral (D-V) and nasal-temporal (N-T) axes. The D-V axis is established by a ventral to dorsal gradient of VAX2, whereas the N-T axis is coordinated by expression of the forkhead transcription factors FOXD1 and FOXG1. Additional gradients are formed within the retina. This spatial distribution may aid in proper targeting of RGC axons that function to establish the retinotopic map.
Blood supply
wikipedia, wiki, encyclopedia, book, article, read, free download, Information about retina. What is retina? What does retina mean?